Молекулы, организмы и естественный отбор

Иногда высказывалось мнение, что успех «Происхождения» только доказывал, что «вопрос уже носился в воздухе» и что «умы были к нему подготовлены». Но я не думаю, чтобы это было верно… Раза два или три пытался я объяснять очень способным людям, что разумею под «естественным отбором», но совершенно безуспешно. Безусловно верно, я думаю, только то, что в умах натуралистов уже успели накопиться бесчисленные и точно наблюдаемые факты, ожидавшие только ясной теории, в которой они нашли бы себе надлежащее место.
Чарльз Дарвин
 |
|
Чарльз Дарвин |
В середине XIX века два ученых, работавших в разных странах и в разных областях естественных наук, почти одновременно сделали два открытия, которые в тот момент выглядели совершенно не связанными между собой.
Первым из этих ученых был всем известный Чарльз Дарвин. В 1859 году вышло первое издание его книги «Происхождение видов путем естественного отбора», где был описан вероятный механизм биологической эволюции. Началось победное шествие дарвиновской теории.
Через два года, в 1861 году, русский химик Александр Михайлович Бутлеров опубликовал статью под скромным названием «К истории производных метилена». Он обнаружил, что нагревание формальдегида в щелочной среде приводит к синтезу сахаристых веществ. Реакция Бутлерова уже дважды обсуждалась в журнале «Химия и жизнь» — в статье В.Н. Пармона «Новое в теории появления жизни» (2005, № 5) и в статье М.А. Никитина «Проблема хиральной чистоты» (2013, № 3), поэтому здесь мы обойдемся без химических подробностей.
Ни Дарвин, ни Бутлеров, скорее всего, и не подозревали, что подошли с разных сторон к исследованию одного и того же чрезвычайно масштабного явления. Общепринятый термин для обозначения этого явления появился только через сто с лишним лет. В 1976 году вышла книга Ричарда Докинза «Эгоистичный ген», в которой автор ввел понятие репликатора.
Репликатором называется молекула, способная в подходящей среде создавать копии самой себя. Иногда понятие репликатора расширяют, охватывая не только молекулы, но и куда более сложные структуры. Но принцип всегда один: репликатор — это объект, так или иначе содействующий созданию собственных копий. Попросту говоря, репликатор «умеет» размножаться.
Теорию репликаторов и их участия в биологической эволюции основательно разработали английский биолог Джон Мейнард Смит (John Maynard Smith) и венгерский биолог Эрш Сатмари (Eörs Szathmáry). Профессор Мейнард Смит умер в 2004 году, а профессор Сатмари продолжает свое творчество и сейчас. Его идеи вполне можно охарактеризовать как современную интерпретацию дарвинизма. На них и будет основана первая часть этой статьи.
 |
|
А.М. Бутлеров |
От катализа к эволюции
Самые простые репликаторы, какие существуют в природе, можно найти в автокаталитических химических реакциях. Что значит «автокаталитические»? Химическая реакция — это превращение одних веществ в другие. Ясно, что химические реакции могут идти с разными скоростями. Вещество, которое ускоряет реакцию, но само выходит из нее, не изменившись, называется катализатором.
Допустим, например, что превращение вещества X в вещество Y катализируется веществом A. Это будет обычная каталитическая реакция:
A + X A + Y
Некоторые катализаторы «умеют» заодно катализировать синтез самих себя. Допустим, что в нашей реакции на входе фигурирует по одной молекуле веществ X и A, а на выходе получается молекула вещества Y и две молекулы вещества A:
A + X 2A + Y
Вот такая реакция и называется автокаталитической. Принимающее в ней участие вещество A можно считать простейшим репликатором. Оно «размножается».
Первую известную химикам автокаталитическую реакцию открыл А.М. Бутлеров. Реакция Бутлерова относится к органической химии, то есть к химии соединений углерода. Исходным веществом, с которого Бутлеров начал, был формальдегид. Его молекула содержит всего один атом углерода. В условиях, созданных Бутлеровым, из формальдегида рано или поздно синтезировался гликольальдегид — более сложное вещество, в молекуле которого атомов углерода уже два. После чего гликольальдегид начинал катализировать синтез еще более сложных сахаров… и самого себя. Молекула гликольальдегида становилась репликатором.
Следующий логический шаг состоит в том, что репликатор может воспроизводить себя абсолютно точно, а может и с вариациями. Допустим, что в нашей воображаемой реакции вместо двух одинаковых молекул катализатора A получается одна молекула A и одна слегка отличающаяся от нее молекула A-штрих:
A + X A + A’ + Y
Тогда новая версия репликатора может вытеснить старую. Начнется эволюция — если не биологическая, то химическая. Химические системы тоже могут эволюционировать по Дарвину. Самая настоящая дарвиновская эволюция, да еще и очень бурная, наблюдается в искусственной бесклеточной среде, где «живут» воспроизводящие себя из поколения в поколение молекулы РНК. Авторы этих экспериментов так сформулировали вопрос, на который хотели получить ответ: «Что произойдет с молекулами РНК, если единственным предъявленным к ним требованием будет библейский призыв ”размножайтесь!”, дополненный биологическим условием делать это как можно быстрее?» Результаты оказались крайне интересными (если вы о них еще не слышали, погуглите по словосочетанию «монстры Шпигельмана»). Трудно представить более наглядную иллюстрацию работоспособности дарвиновского эволюционного механизма. Впрочем, тут мы забегаем вперед.
В реакции Бутлерова участвует не только гликоль-альдегид. Кроме него роль автокатализаторов там могут играть глицеральдегид, дигидроксиацетон и некоторые другие органические молекулы. Но состав конечных продуктов — сложных сахаров — в этой реакции всегда одинаков, от состава катализаторов он не зависит. Эта система, грубо говоря, не дотягивает до эволюции, потому что в ней нет обратной связи, позволяющей конечному результату влиять на начальные условия.
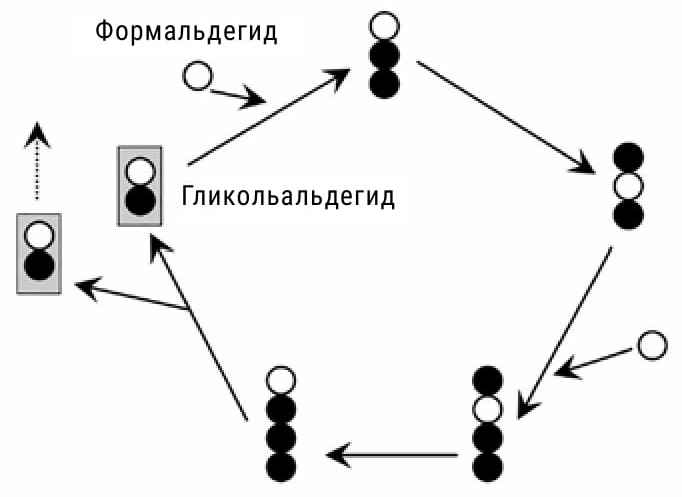 |
|
Реакция Бутлерова без химических формул. Каждый кружочек обозначает атом углерода (белый кружочек — атом, принадлежащий формальдегиду). Гликольальдегид вступает в реакцию, которая после цикла превращений приводит к синтезу еще одной молекулы гликольальдегида. |
Ограниченная наследственность
Эрш Сатмари делит все возможные репликаторы на две группы — холистические и модульные. Холистический репликатор целостен: он воспроизводится по принципу «все или ничего». К таким репликаторам относится гликольальдегид в реакции Бутлерова. Модульный репликатор состоит из частей, которые в той или иной мере независимы друг от друга и могут меняться по отдельности. Такой репликатор может копировать себя точно, а может и с вариациями. Важным частным случаем модульного репликатора является матричный репликатор, который копирует текст, то есть линейную последовательность знаков.
Уместно добавить, что гипотезу о существовании матричных репликаторов — без этих терминов, но абсолютно современную по сути — впервые высказал в 1927 году Николай Константинович Кольцов, 150-летие со дня рождения которого мы отметили совсем недавно. «Химия и жизнь» не раз писала о творчестве Кольцова (см.1965, № 5; 1972, № 7; 2001, № 7 и 2018, № 12).
Кроме того, Сатмари вводит понятие репликаторов с ограниченной и неограниченной наследственностью. Что это значит?
Рассмотрим очень простой матричный репликатор, который получил в 1986 году немецкий биохимик Гюнтер фон Кедровски (Günter von Kiedrowski). Он представляет собой гексануклеотид, то есть цепочку из шести нуклеотидов. Как известно, нуклеотиды в составе ДНК бывают четырех типов: адениновый, тиминовый, гуаниновый и цитозиновый, сокращенно А, Т, Г и Ц. Нуклеотидная последовательность репликатора фон Кедровски — ЦЦГЦГГ. Оказалось, что при некоторых условиях такая молекула может успешно воспроизводить себя, причем без всякой помощи обычно присутствующих в живой клетке ферментов.
Наследственность — это способность передавать из поколения в поколение индивидуальные различия. У матричных репликаторов, состоящих из нуклеиновых кислот, эти различия выражаются в последовательности нуклеотидов. Поставим вопрос: сколько вариантов нуклеотидной последовательности максимально возможно для репликатора фон Кедровски?
Ответить на это нетрудно. Репликатор фон Кедровски — это цепочка из шести нуклеотидов. Допустим, что на каждой из шести позиций в этой цепочке может находиться нуклеотид любого из четырех типов: А, Т, Г или Ц (на самом деле это не так — в опыте фон Кедровски на нуклеотидную последовательность были наложены жесткие ограничения, но мысленный эксперимент дает нам право их проигнорировать). Тогда общее число потенциально возможных вариантов репликатора будет равно 46 = 4096. По меркам химии это очень маленькое число. Оно намного меньше числа молекул в любой реалистичной химической системе, вроде колбы или пробирки с раствором. А значит, с размножением репликатора все его варианты очень скоро будут перебраны. Это и есть репликатор с ограниченной наследственностью: число его возможных вариантов меньше численности репликатора в тех условиях, в которых он «живет».
Темная геномная Вселенная
В качестве более сложного репликатора представим себе отрезок ДНК, который кодирует белок, состоящий из 33 аминокислот (для настоящего белка это очень мало, но не будем придираться). Генетический код у земных организмов триплетный: каждая аминокислота в белке кодируется тремя нуклеотидами. Значит, участок ДНК, кодирующий наш белок, должен состоять из 99 пар нуклеотидов.
Сколько вариантов может иметь такая последовательность? Ответ: 499 = 1060. Это астрономическое число. По порядку величины оно соответствует примерному времени жизни Вселенной, если его измерять планковскими временами, то есть наименьшими интервалами времени, у которых есть физический смысл. Планковское время, оно же квант времени, имеет величину 5,4∙10−44 cекунды. С другой стороны, синтез белка — процесс довольно медленный, его длительность измеряется целыми секундами, а то и минутами. Вероятно, даже за миллиарды лет перебрать все варианты нашего белка эволюция не успела бы.
Это далеко не предел. Геном человека состоит примерно из 3 миллиардов пар нуклеотидов (точная величина, если не учитывать дублирование генетической информации, равна 3,1∙109). Тогда полное число вариантов последовательности размером с геном человека будет равно четверке в степени три миллиарда. Это — уже не астрономическое число. В обычной природе подобными числами просто нечего мерить. Единственная область естествознания, где они встречаются, — это инфляционная космология, раздел теоретической физики, рассматривающий гипотезы о стремительном раздувании Вселенной в самом начале ее истории.
«Представим себе пространство, каждая точка которого соответствует одному варианту генома человека. Изначально оно абсолютно темное. Секвенируем все существующие индивидуальные геномы и подсветим точку, соответствующую каждому из них. Пространство все равно останется почти полностью темным», — пишет Эрш Сатмари. В этом смысле эволюция жизни только началась.
На грани жизни
У холистических репликаторов наследственность всегда ограниченна (во всяком случае, никаких обратных примеров мы не знаем). Модульные и, в частности, матричные репликаторы могут обладать как ограниченной, так и неограниченной наследственностью. Но перейти порог, отделяющий ограниченную наследственность от неограниченной, матричным репликаторам очень легко: он проходит где-то на уровне цепочки из нескольких десятков нуклеотидов. Все современные природные генетические системы находятся далеко за этим порогом. Гены, хромосомы, геномы — это образцовые матричные репликаторы с неограниченной наследственностью. Вот они-то и имеют самый большой потенциал для эволюции. Механизм этой эволюции, открытый Чарльзом Дарвином, называется естественным отбором.
Надо подчеркнуть, что Дарвин ничего не знал ни о репликаторах, ни о цифровом характере наследственной информации. Сам он, выражаясь современным языком, считал ее аналоговой. Эту простительную ошибку удалось исправить только через сто лет, когда был расшифрован генетический код. В сущности, любой матричный репликатор — это цифровой репликатор: информация в нем кодируется набором единиц, которые отличаются друг от друга четко, как буквы алфавита. В генетических репликаторах такими единицами служат, естественно, нуклеотиды. Дарвин не имел обо всем этом ни малейшего представления, но это не уменьшает, а увеличивает его заслугу. До возникновения генетики, которая могла бы ответить на многие из волновавших его вопросов, он просто не дожил.
Джон Мейнард Смит и Эрш Сатмари утверждают, что мы знаем две — всего лишь две! — природные системы репликаторов, имеющих неограниченную наследственность. Первая из них — это гены и геномы, а вторая — человеческий язык.
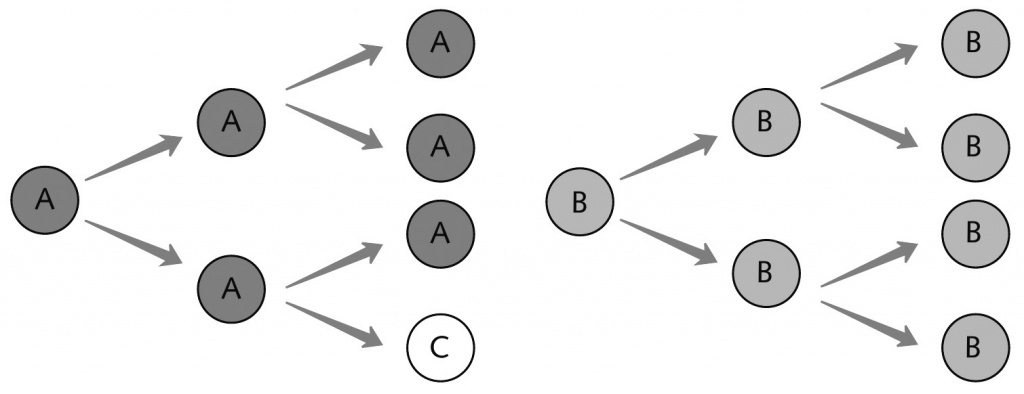 |
|
Эту картинку Джон Мейнард Смит и Эрш Сатмари называют определяющей характеристикой жизни. Особи с разными генотипами (A, B) размножаются, при этом в результате мутации иногда может возникнуть новый генотип (C), и все они начинают конкурировать за ресурсы. |
Главная тайна Дарвина
Теперь мы можем сформулировать определение естественного отбора. Итак, естественный отбор — это неслучайное воспроизведение генотипов (в отличие от случайного, которое называется дрейфом генов).
Поясним это определение. У любого живого организма есть геном, который служит в ряду поколений репликатором. Он состоит из репликаторов более низкого порядка, но тоже матричных — таких, как отдельные гены. Геном включает в себя всю генетическую информацию, какая есть у данного биологического вида. Однако известно, что варианты (аллели) многих генов, входящих в геном, у разных особей одного и того же вида могут отличаться. Поэтому любая отдельная особь может обладать своим собственным вариантом генома — генотипом. На отвлеченных схемах генотипы можно для простоты обозначать буквами — A, B, C и так далее.
Когда особи размножаются, генотипы копируются. Но это копирование всегда неточно: случайная мутация может превратить один генотип в другой (например, генотип A в генотип C). Генотипы, оказавшиеся «неудачными», исключаются из преемственности поколений — как говорят биологи, элиминируются (это не значит, что их носители непременно погибают: чтобы лишиться шанса передать свои гены потомкам, достаточно не участвовать в размножении). Но и относительно «удачные» генотипы, как правило, обладают разными свойствами, а потому воспроизводятся с разными скоростями. В условиях ограниченных ресурсов (а в природе ресурсы ограничены всегда) это приводит к тому, что одни генотипы со временем вытесняют другие. Генотипический состав популяции меняется. Это и есть дарвиновская эволюция.
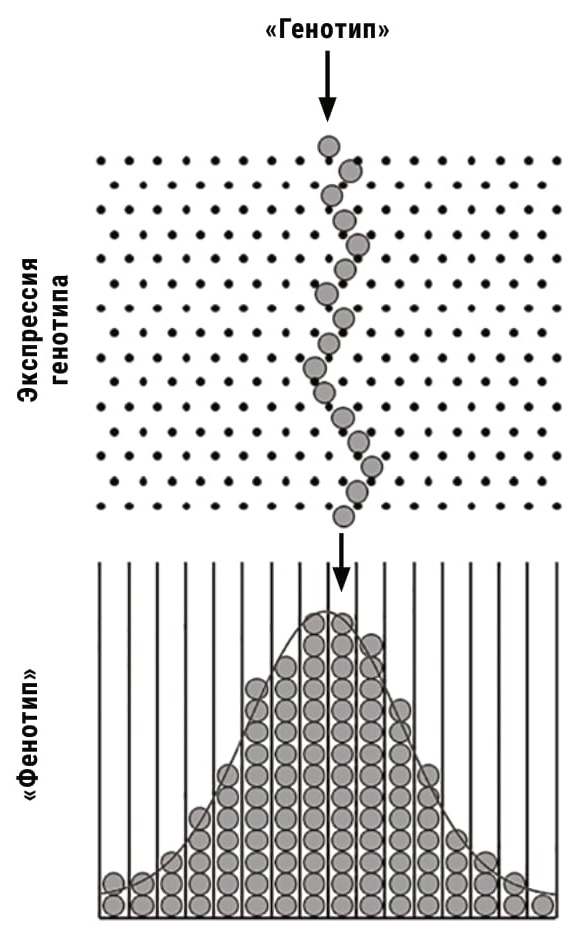 |
|
Генотип, сталкиваясь с внешними и внутренними помехами, порождает разнообразие фенотипов подобно тому, как падающие дробинки в аппарате Гальтона, сталкиваясь с булавками, распределяются по разным отделениям ящика. Это распределение описывается гауссовой кривой. |
Генотип и фенотип
Настоящая сложность состоит в том, что генотипы отбираются не напрямую, а через посредство фенотипов. Генотип — это в конечном счете просто набор нуклеотидов. Сам по себе он бесполезен, как книга, у которой нет читателей. «Читатель» генотипа — это живая клетка, молекулярная машинерия которой умеет синтезировать белки (а также РНК) на основе инструкций, заключенных в нуклеотидном тексте. Будучи синтезированы, белки делают свою работу: катализируют в клетке химические реакции, встраиваются в мембрану, преобразуют энергию, переносят те или иные молекулы, придают частям клетки подвижность, меняют ее форму, организуют деление и выполняют еще массу функций. Сумма признаков, которые приобретет клетка в результате, и будет фенотипом, соответствующим ее генотипу. Ну а у многоклеточных организмов одни и те же гены в разных клетках работают по-разному, и вся система становится еще более сложной.
У простых репликаторов, которые еще нельзя считать живыми существами, дистанция между «генотипом» и «фенотипом» очень мала. Например, у воспроизводящихся в пробирке молекул РНК она сводится к сворачиванию нуклеотидной цепочки, которое целиком диктуется законами химии. У вирусов эта дистанция уже больше, у одноклеточных организмов — намного больше. А у многоклеточных животных и растений она просто колоссальна.
Воспользуемся моделью, которую придумал двоюродный брат Чарльза Дарвина — английский ученый Фрэнсис Гальтон (Francis Galton). Глубокого научного смысла в ней нет, зато она очень наглядна. Представим себе вертикально поставленный плоский ящик, в который через узкую воронку падают шарики. Низ ящика разгорожен на ряд одинаковых отделений. Если бы шарики падали свободно, они все попали бы в центральное отделение, расположенное прямо под воронкой. Но в стенку ящика вбиты булавки, которые не дают шарикам падать по прямой: сталкиваясь с булавками, они случайно отклоняются то в одну, то в другую сторону.
В центральное отделение, конечно, в любом случае попадет больше всего шариков, но далеко не все. А чем дальше отделение от центра, тем меньше шариков до него долетит. Если аккуратно провести этот опыт, распределение шариков по отделениям будет довольно точно описываться колоколообразной кривой, носящей имя Карла Фридриха Гаусса. (О Гальтоне и его модели см. «Химию и жизнь», 2022, № 3.)
В этой модели источник шариков соответствует генотипу. Булавки — это всевозможные помехи на пути реализации генетической программы. Ну, а разброс шариков по отделениям — это полное множество фенотипов, возможных при данном генотипе (или группе генотипов), которое принято называть нормой реакции.
Формы отбора
Теоретики эволюционной биологии выделили десятки разных форм естественного отбора. Их полный обзор — задача учебников, а не популярного очерка. Но есть как минимум две формы отбора, не зная которых обсуждать эволюцию почти невозможно. Это — стабилизирующий и движущий отбор.
Стабилизирующий отбор — это самая универсальная форма естественного отбора. Его существование вытекает из предельно общих свойств жизни. Мы знаем, что генотипы никогда не копируются абсолютно точно: в каждом поколении появляются новые мутации. Давление мутаций стремится расширить норму реакции, растягивая гауссовский «колокол» по горизонтали. Возникают фенотипы, далекие от стандартных, — как правило, не очень-то жизнеспособные. Естественный отбор устраняет эти фенотипы вместе с определяющими их генами, возвращая норму реакции на место. Вот такой отбор и называется стабилизирующим. Он действует почти постоянно, и только благодаря ему каждый биологический вид сохраняет свой неповторимый облик.
Если уклонения от стандартного фенотипа резки и дискретны, мы называем их уродствами. Каждому ясно, что уродства обычно не способствуют выживанию. Но стабилизирующий отбор может действовать и намного тоньше. Часто он отсекает не только явные уродства, но и едва заметные количественные отличия. Например, у растений, опыляемых насекомыми, стабилизирующий отбор строго контролирует пропорции цветка: чтобы хоботок насекомого-опылителя дотянулся точно до тычинок, венчик не должен быть ни слишком высоким, ни слишком низким — иначе растение потеряет шанс оставить потомство.
Движущим отбором называется отбор против одного из крайних вариантов признака в пользу другого крайнего варианта. Под давлением движущего отбора норма реакции не расширяется и не сужается — она смещается. Стандартный фенотип становится другим. Конечно, для этого отбор должен быть эффективным и действовать достаточно долго, не меняя направления. Это и есть тот самый процесс, который, по Дарвину, создает новые формы живых организмов.
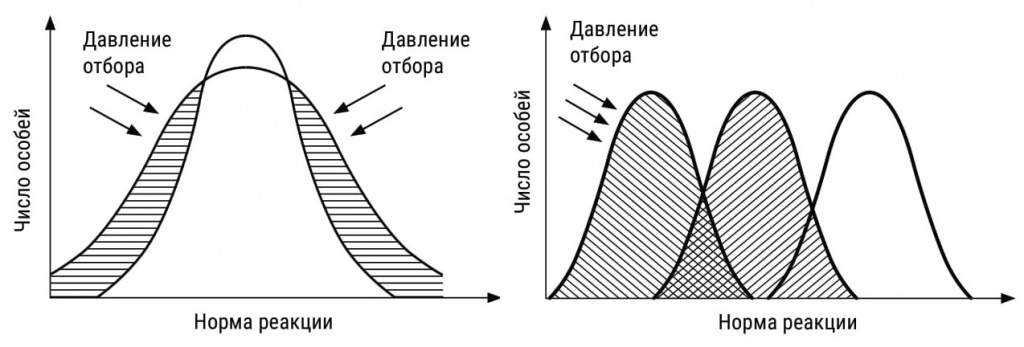 |
|
Стабилизирующий и движущий отбор. Норма реакции, включающая в себя множество всех возможных фенотипов, под давлением стабилизирующего отбора сужается и удерживается на месте, а под давлением движущего — смещается. |
Псевдогены
«Счастье — как здоровье: когда оно налицо, его не замечаешь», — писал М.А. Булгаков в рассказе «Морфий». Если бы великий писатель изучил эволюционную теорию, он наверняка сказал бы, что стабилизирующий отбор подобен здоровью: его трудно заметить, когда он есть, и очень легко — когда его нет.
Представим, что стабилизирующий отбор перестал действовать. Что будет? Ответ: начнется немедленный рост изменчивости по тем генам и признакам, которые раньше контролировались стабилизирующим отбором, а теперь перестали. Этот процесс вполне можно будет наблюдать, в том числе и на уровне нуклеотидных последовательностей.
Вот один пример. В эволюции животных и растений часто происходят события, которые называются дупликациями генов. Термин «дупликация» означает, что ген в буквальном смысле удваивается. Причины дупликаций разнообразны, но сейчас нам не важны. Нам важно, что в результате дупликации в геноме оказывается дополнительная копия гена, который и без того хорошо работает.
После удвоения гена мутации, нарушающие работу одной из его копий, перестают быть вредными. Они больше не отсеиваются стабилизирующим отбором — ведь есть вторая копия гена, которой организму вполне хватает. Рано или поздно накапливающиеся мутации захватывают регуляторные участки гена, необходимые для того, чтобы он мог включаться. Тогда ген выходит из строя. Нефункционирующий молчащий ген называется псевдогеном. Псевдогенизация — это «смерть» гена. И наступает она именно благодаря ослаблению стабилизирующего отбора.
На уже «мертвые» псевдогены стабилизирующий отбор не действует, поэтому, как и следовало ожидать, их нуклеотидные последовательности меняются в ходе эволюции в несколько раз быстрее, чем у функционирующих генов. Тем не менее псевдогены могут сохраняться в геномах очень долго. Это — важные свидетельства древних эволюционных событий.
«Лишние» гены, возникшие в результате дупликаций, превращаются в псевдогены часто, но не всегда. Ослабление стабилизирующего отбора дает гену на короткое время огромную эволюционную свободу, открывающую окно возможностей. Бывает, что в разных копиях гена закрепляются разные мутации, и между ними происходит разделение функций. А бывает и так, что одна из копий гена получает в итоге совершенно новую функцию. В эволюции встречаются целые серии последовательных дупликаций, после которых гены делят между собой функции или приобретают новые. Группа генов, происходящих таким путем от одного гена-предка, называется семейством генов.
Например, у большинства многоклеточных животных есть семейство генов, которое называется Hox. Эти гены важны для дифференцировки отделов тела и для развития нервной системы. У позвоночных Hox-гены сгруппированы в несколько кластеров, находящихся в разных хромосомах. Исходно в каждом кластере было по 14 генов, но часть из них в ходе эволюции исчезла. Гены, которые называются Hoxa14 и Hoxb14, отсутствуют в двух самых крупных эволюционных ветвях — у наземных позвоночных и у лучеперых рыб. У хрящевых рыб — акул — эти два гена сохранились, но в псевдогенизированном состоянии. Это типичный случай превращения в псевдогены генов, которые не очень нужны. А вот у единственной современной кистеперой рыбы латимерии ген Hoxa14 до сих пор остается функциональным. И у австралийской двоякодышащей рыбы рогозуба — тоже. Тут мы видим большую архаичность этих рыб, близких к предкам наземных позвоночных, и одновременно — свидетельство их особого эволюционного пути.
Жил на свете таракан
Движущий отбор легче всего наблюдать у организмов, находящихся под антропогенным давлением. В этих случаях среда меняется очень быстро, и организмы волей-неволей должны успевать к ней приспосабливаться. Внимание: мы сейчас говорим не об искусственном отборе, когда человек намеренно отбирает нужные ему формы, а о естественном, когда он действует как слепой природный фактор, ничем не хуже и не лучше извержения вулкана или эпидемии.
Рыжий таракан (Blattella germanica) обитает в человеческих жилищах по всему миру, от Аляски до Антарктики включительно. Люди его не любят и периодически травят химикатами. Но таракан не сдается. Примерно с 1980-х годов против рыжего таракана стали часто применять отравленные приманки, которые вместе с ядом содержали глюкозу или фруктозу. Эти сладкие для нас вещества привлекают и тараканов, поэтому сначала отравленные приманки действовали. Но уже через несколько лет у тараканов выработалось наследственное отвращение к глюкозе, быстро распространившееся по их популяциям. Как это могло произойти?
Дело в том, что у таракана прекрасно развито чувство вкуса. В придатках ротового аппарата у него, как и у других насекомых, сидят вкусовые рецепторные нейроны, каждый из которых специализирован на определенном типе веществ. В клеточные мембраны этих нейронов встроены рецепторы — белки, распознающие окружающие молекулы по известному в биохимии принципу ключа и замка. Есть рецепторы сахаров («сладкий вкус»), а есть рецепторы широкого спектра бесполезных или ядовитых веществ («горький вкус»). Каждый белок-рецептор, как и вообще любой белок, кодируется своим геном. При этом нейронные сети в мозгу таракана устроены так, что рецепторы «горького вкуса» имеют приоритет: они заставляют таракана избегать пищи, даже если рецепторы «сладкого вкуса» в этот момент тоже активны.
В данном случае у таракана произошла одна-единственная мутация. Она изменила молекулярную структуру рецептора «горького вкуса» таким образом, что он стал реагировать не только на яды, но и на глюкозу. В результате нервная система таракана стала воспринимать глюкозу как яд. Подобные мутации наверняка происходили и раньше, но в обычных условиях они были вредными. И действительно, показано, что тараканы, которые шарахаются от глюкозы, медленнее растут. Но в окружении отравленных приманок, где привлекающим фактором служила именно глюкоза, эта мутация оказалась спасительной. Движущий отбор тут же ее подхватил, и обладатели отвращения к глюкозе стали составлять большинство популяции.
Мыши на лаве
История с тараканами показывает нам удачный пример поиска мишени — гена или белка, на который в конечном счете действует движущий отбор. Рассмотрим еще один пример, более сложный, зато и более интересный.
В юго-западной части США (штаты Аризона, Нью-Мексико) и на севере Мексики живет мышевидный грызун под названием скальный щетинистый прыгун (Chaetodipus intermedius). Главная особенность этого зверька ясна из его названия: он — петрофил, то есть живет на скалах. Среди грызунов такие виды попадаются.
Зоологи давно заметили, что разные популяции скального прыгуна сильно отличаются друг от друга окраской шерсти. Дело в том, что скальные выходы в тех местах бывают разных цветов. На светлых каменистых россыпях живут прыгуны, шерстка которых тоже светлая, песчаного цвета. Таких популяций большинство. Но на темной застывшей базальтовой лаве они другие — почти черные. Приспособительный смысл тут ясен: окраска помогает зверькам скрываться от хищников, особенно от сов (щетинистые прыгуны ведут ночной образ жизни). Но как именно эта адаптация была достигнута?
Окраска млекопитающих определяется пигментом меланином, который синтезируется в особых клетках — меланоцитах. У этих клеток есть тонкие древовидные отростки, куда транспортируется готовый меланин, упакованный в компактные гранулы — меланосомы. Из отростков меланоцитов меланосомы переходят в клетки, формирующие волосы. Меланин бывает разных типов: черно-коричневый — эумеланин или желто-красный — феомеланин. Окраска кожи и волос зависит от их соотношения.
Работу меланоцитов регулирует гормон гипофиза, который называется меланоцитстимулирующим гормоном, сокращенно МСГ. Чем сильнее действует этот гормон, тем больше эумеланина вырабатывается в организме. Но что значит «сильнее действует»? Чтобы подействовать, молекула гормона должна связаться с рецептором — специальным белком, который сидит в мембране клетки-меланоцита и передает внутрь клетки полученный сигнал. Эффект гормона зависит не только от его концентрации в крови, но и от того, насколько сильно рецептор на него реагирует.
А теперь — главное. В интересующем нас случае рецептором МСГ служит мембранный белок под названием MC1R. Мутации в гене, кодирующем этот белок, могут слегка изменить его структуру таким образом, что он будет либо прочнее связывать молекулу МСГ, либо интенсивнее передавать сигнал внутрь клетки. И то и другое усилит синтез эумеланина. Именно такие мутации и обнаружились в меланистических (то есть темноокрашенных) популяциях скального прыгуна. И что особенно интересно, в разных меланистических популяциях эти мутации — разные. Хотя фенотипический эффект они дают один и тот же: потемнение шерсти. Дело в том, что поля застывшей базальтовой лавы в ареале скального прыгуна довольно редки и живущие на них популяции изолированы друг от друга. Темный вариант окраски выработался у них независимо: в разных случаях движущий отбор подхватил разные мутации, действующие на один и тот же белок.
Судя по всему, эта адаптация может при нужде возникнуть очень быстро. Одно из лавовых полей, заселенных скальными прыгунами, имеет возраст всего в 1000 лет — очень небольшой срок по эволюционным меркам! — но живущая там популяция уже успела стать меланистической. Скорости отбора для этого хватило.
Почему неправ Берг?
Дальнейшие исследования показали, что мутации, действующие на белок MC1R, приводят к потемнению окраски не только у скального щетинистого прыгуна, но и у многих других млекопитающих и даже у птиц. Как это работает?
Белок MC1R представляет собой цепочку из примерно трехсот аминокислот. У разных видов птиц и млекопитающих (включая, между прочим, и человека) удалось идентифицировать 16 различных аминокислотных замен в этом белке, которые все приводят к одному и тому же эффекту: рецептор MC1R начинает сильнее реагировать на сигнал, переданный молекулой МСГ. С точки зрения биофизики достичь такого результата не труднее, чем вызвать короткое замыкание в электросети. В итоге синтез эумеланина усиливается, и окраска темнеет.
Тут-то мы и выходим на самые общие проблемы биологии. В 1922 году ихтиолог и географ Лев Семенович Берг выпустил книгу «Номогенез, или Эволюция на основе закономерностей». Берг считал, что наследственных вариаций, реально существующих в природе, для дарвиновского естественного отбора просто не хватит. Он не спорил с тем, что эволюция путем естественного отбора логически мыслима. Но он был уверен, что теория Дарвина не выдерживает проверки фактами. Безграничной изменчивости, которая (по мнению Берга) нужна для естественного отбора, не наблюдается ни в ископаемой, ни в современной природе. Организмы слишком устойчивы, чтобы допустить ее. «Случайный новый признак очень легко может испортить сложный механизм, но ожидать, что он его усовершенствует, было бы в высшей степени неблагоразумно», — пишет Берг. И делает вывод: «Отбору не из чего выбирать».
История белка MC1R (а в современной научной литературе таких историй тысячи) показывает, что Берг был неправ. Изменчивость, конечно, не безгранична. Но ведь все, что требовалось в нашем случае, — это чтобы белковый рецептор слегка «заедало» во включенном положении. Мы видим, что мутаций, делающих это сравнительно безопасно для организма, не одна и не две. Сейчас, когда мы добрались до молекулярных мишеней отбора, можно точно сказать, что тезис Берга неверен. Отбору очень даже есть из чего выбирать. И на уровне функциональных белков это, скорее, правило, чем исключение.
Красное и черное
Иногда движущий отбор остается незавершенным. Например, на маленьких островках у побережья Шотландии живут одичавшие овцы, самцы которых отличаются друг от друга размером рогов. У одних баранов рога большие, у других — маленькие, зачаточные. Большерогие бараны успешнее размножаются, но хуже выживают зимой. Поэтому отбор идет одновременно в разные стороны и сохраняет более одного генотипа. Как говорят генетики, между векторами отбора установилось равновесие. В результате в единой популяции присутствуют две резко различные формы. Прерывистое разнообразие форм внутри вида, не связанное ни с какими барьерами для скрещивания, называется полиморфизмом.
Классический пример полиморфизма описал Николай Владимирович Тимофеев-Ресовский у двухточечной божьей коровки (Adalia bipunctata). У этого вида жуков очень изменчива окраска. Разные цветовые формы отличаются друг от друга соотношением красного и черного. Многолетние наблюдения показали, что «черные» божьи коровки успешнее размножаются летом, зато «красные» лучше переносят зимовку. Возникает циклический отбор, благоприятствующий то одному генотипу, то другому. В результате в популяциях все время есть и «черные», и «красные» божьи коровки: осенью — примерно 60% «черных» и 40% «красных», а весной соотношение меняется на обратное.
Добавим, что Тимофеев-Ресовский проводил свои наблюдения в окрестностях Берлина, где двухточечная божья коровка, как правило, дает в теплый сезон три поколения. В более северных странах она дает только одно поколение. Тогда преимущество «черной» формы в скорости размножения не успевает реализоваться. Но полиморфизм по окраске в этих странах все равно есть. Так что, видимо, на него влияют и другие факторы. Как обычно, в природе все сложнее, чем в моделях.
Одной из причин полиморфизма — не единственной, но частой — бывает особая форма естественного отбора, которую называют дизруптивным отбором. Мы знаем, что обычный движущий отбор действует против одного из крайних вариантов фенотипа в пользу другого крайнего варианта. А теперь вообразим, что оба крайних варианта (или большее их число) получили преимущество в выживании. Отбор пойдет в пользу этих вариантов и против «золотой середины», занятой промежуточными значениями признака. Вот такой отбор и называется дизруптивным. Под давлением дизруптивного отбора норма реакции разрывается.
Формирующая сила
Можно ли сказать, что естественный отбор служит движущей силой эволюции?
Для начала разберемся, что такое движущая сила. Например, какова движущая сила химических реакций? Это хорошо известно. Движущей силой любой химической реакции служит рассеивание энергии — превращение упорядоченной свободной энергии в беспорядочную тепловую. Ключевое слово тут «энергия». Мы называем движущей силой процесса превращение энергии, благодаря которому этот процесс идет. Например, представим себе движущийся автомобиль. Если это автомобиль с двигателем внутреннего сгорания, то движущей силой служит превращение энергии связей молекул углеводородов (из которых состоит бензин) в тепловую энергию, которая рассеивается в пространстве.
Но к естественному отбору такая интерпретация неприменима. Это процесс, идущий без всяких затрат энергии, «сам собой». Спрашивается, как так может быть?
Вот грубая аналогия, которую предложил больше ста лет назад физик Николай Алексеевич Умов:
«Представим себе органную трубу среди бушующей воздушной бури. Она издает свойственный ей музыкальный тон. На музыкальный тон не было затрачено новой энергии, а только произошел отбор, сортировка хаотических движений частиц воздуха в гармонические или стройные движения. Энергия осталась та же — энергия бури».
Что же служит аналогом «энергии бури» в случае с живыми организмами? Очевидно, это та энергия, которую они всевозможными способами извлекают из внешней среды и превращают в биохимическую энергию своих тел, чтобы затем воспроизводить себя, умножая свою численность по взлетающей вверх экспоненциальной кривой. Потенциальную мощь этого процесса трудно даже вообразить. «Благодаря размножению каждое живое вещество может создать новые любые количества живой материи», — писал по этому поводу Владимир Иванович Вернадский и приводил расчеты: при условии неограниченного размножения холерный вибрион и кишечная палочка могут дать массу живого вещества, равную массе всей земной коры, за время меньше двух суток. «Один из наиболее медленно размножающихся организмов — слон — может дать то же количество вещества в 1300 лет. Но что значат годы и столетия в геологическом, т. е. планетном, времени!»
Этот чудовищный потенциал, основанный на способности живых организмов к неограниченному размножению и на много порядков превосходящий вместимость любой среды, биолог Михаил Михайлович Камшилов назвал давлением жизни. Ясно, что на самом деле размножение неограниченным не бывает. Поверхность Земли вовсе не покрыта сплошь слонами. В реальности выживают далеко не все, кто потенциально мог бы выжить, а это и значит, что происходит отбор.
Итак, движущая сила эволюции — «давление жизни», сталкивающееся с неустранимым противодействием среды. Какова же роль отбора? Это тоже можно пояснить аналогией. У любой реки есть берега. Нельзя сказать, что они — причина течения: вода течет не потому, что она заключена в берегах, а потому что она подчиняется силе тяжести. Тем не менее именно берега, очертания которых диктуются рельефом местности, придают реке форму. Так и естественный отбор — это не движущая, а формирующая сила, virtus formativa в терминологии средневековых философов.
 |
|
Иллюстрация Петра Перевезенцева |
Статья подготовлена по материалам курса лекций об эволюции, который автор читал в 2022 году в культурно-просветительском центре АРХЭ.